
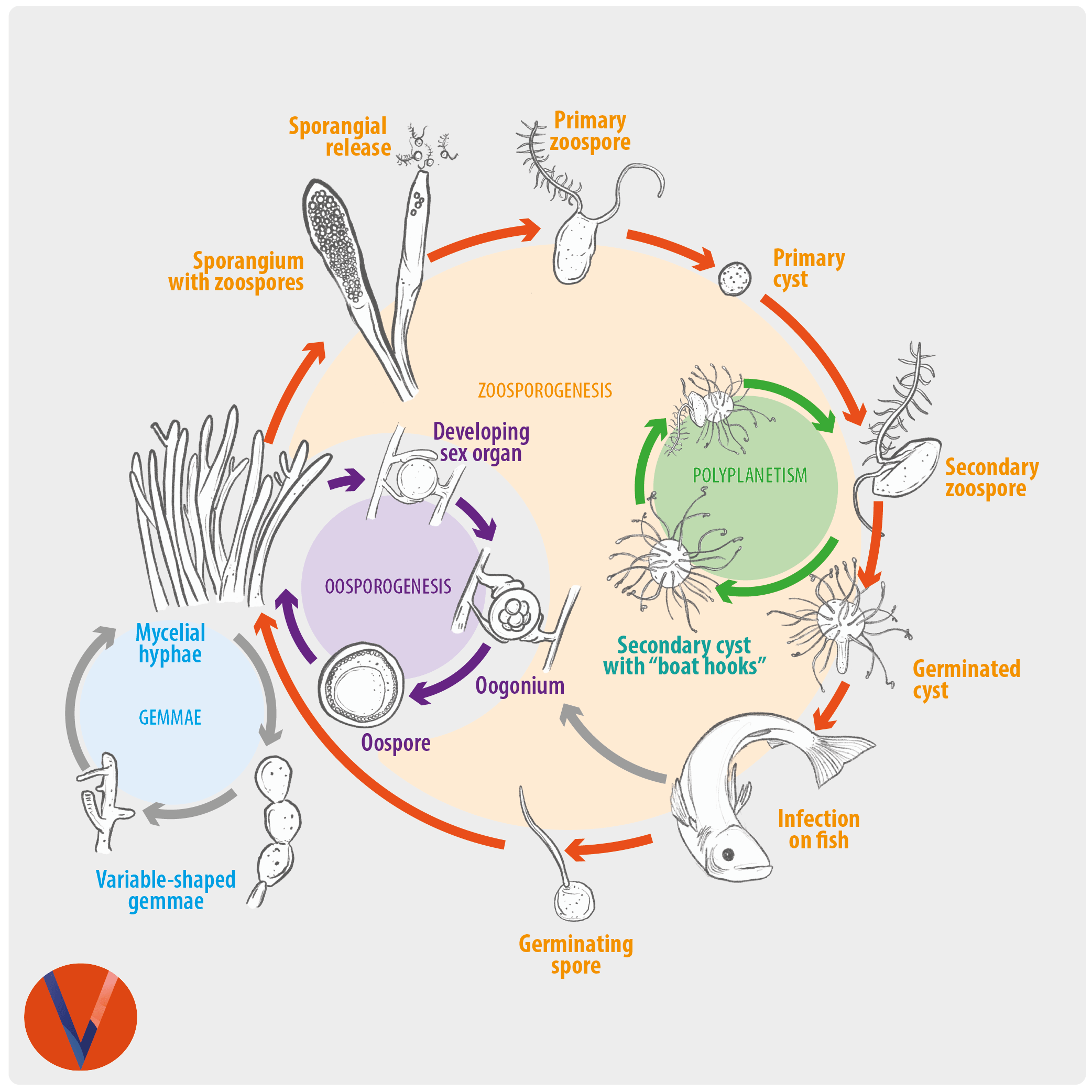
Saprolegnia sp. is classified within Oomycetes, a group of pathogens similar to fungi but which are more closely related to golden-brown algae and are part of the Chromista or chromoalveolates and therefore are not “true fungi”.
Saprolegnia can infect amphibia, molluscs, crustacea, fish and their eggs causing the disease known as Saprolegniasis. The most frequently affected fish are salmonids, which are especially vulnerable to Saprolegnia. Several species of Saprolegnia can be involved as fish pathogens, including Saprolegnia australis, S. parasitica, S. delica, S. diclina, and S. ferax. S. parasitica is considered the most virulent causative agent, leading to devastating infections in freshwater. S. diclina has been considered as the main threat for fish eggs.
Infections with a particularly pathogenic strain may cause huge losses of both fish and eggs. Therefore, specific identification of Saprolegnia strains involved in outbreaks is important in order to decide whether the infection is caused by a pathogenic or saprophytic strain.
Stress factors such as poor water quality, temperature variations, malnutrition, physical injury, reduction in oxygen levels and high density of fish, are well-known risk factors for Saprolegnia infection.
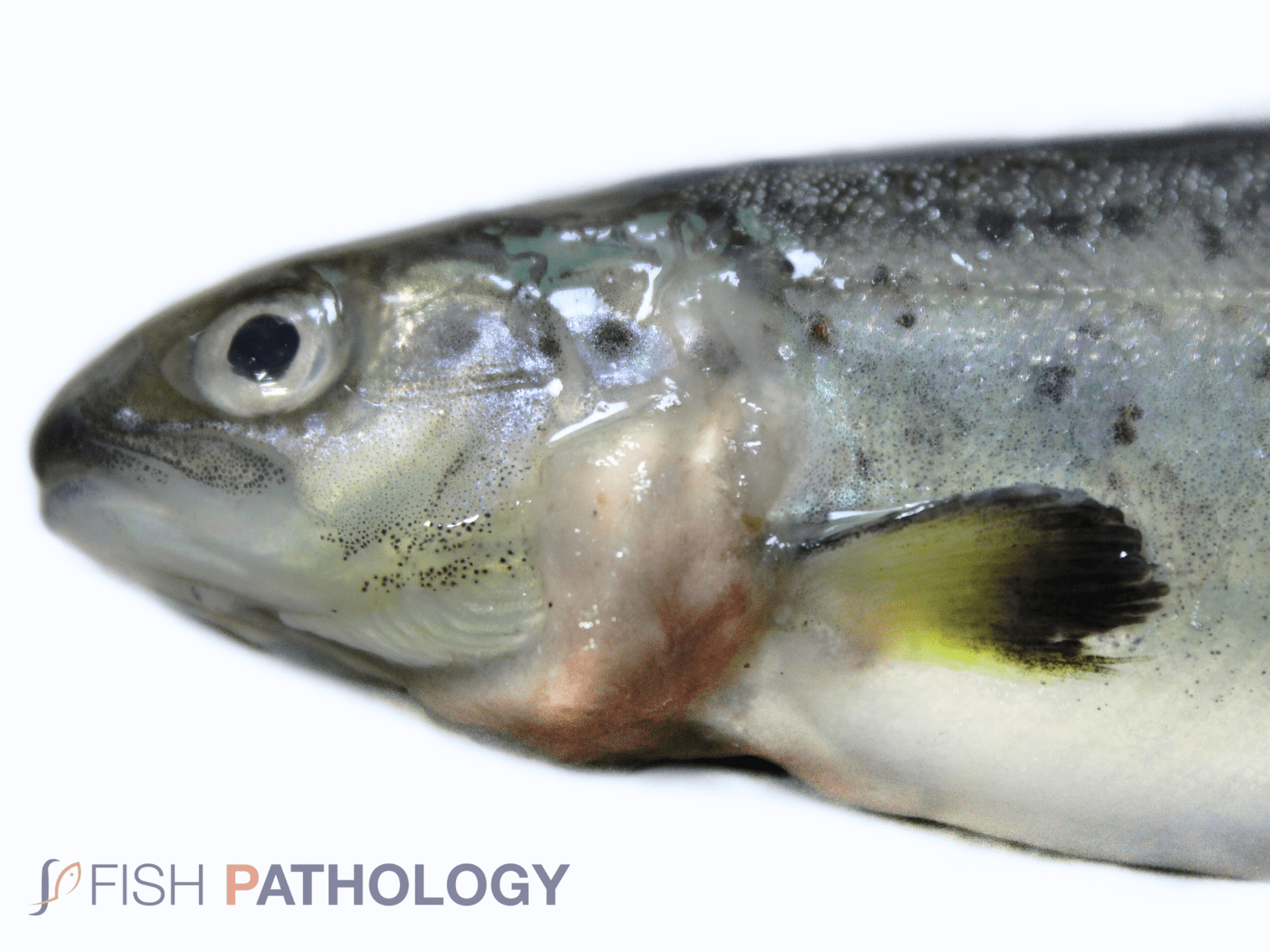
Saprolegnia often presents as a secondary infection that is diagnosed by the appearance of white or grey cotton-like tufts that, when out of water, have a somewhat mucoid appearance. Typical disease signs are visible circular or crescent-shaped, cotton-wool like, white or grey patches of filamentous mycelia on the fish skin. The lesions appear mainly around the head and the caudal, adipose and anal fins. Lesions may spread across the body until adjacent lesions coalesce.
The most virulent strains can penetrate organs causing damage to the underlying muscles and respiratory difficulties may also present when infection is associated with the gills.
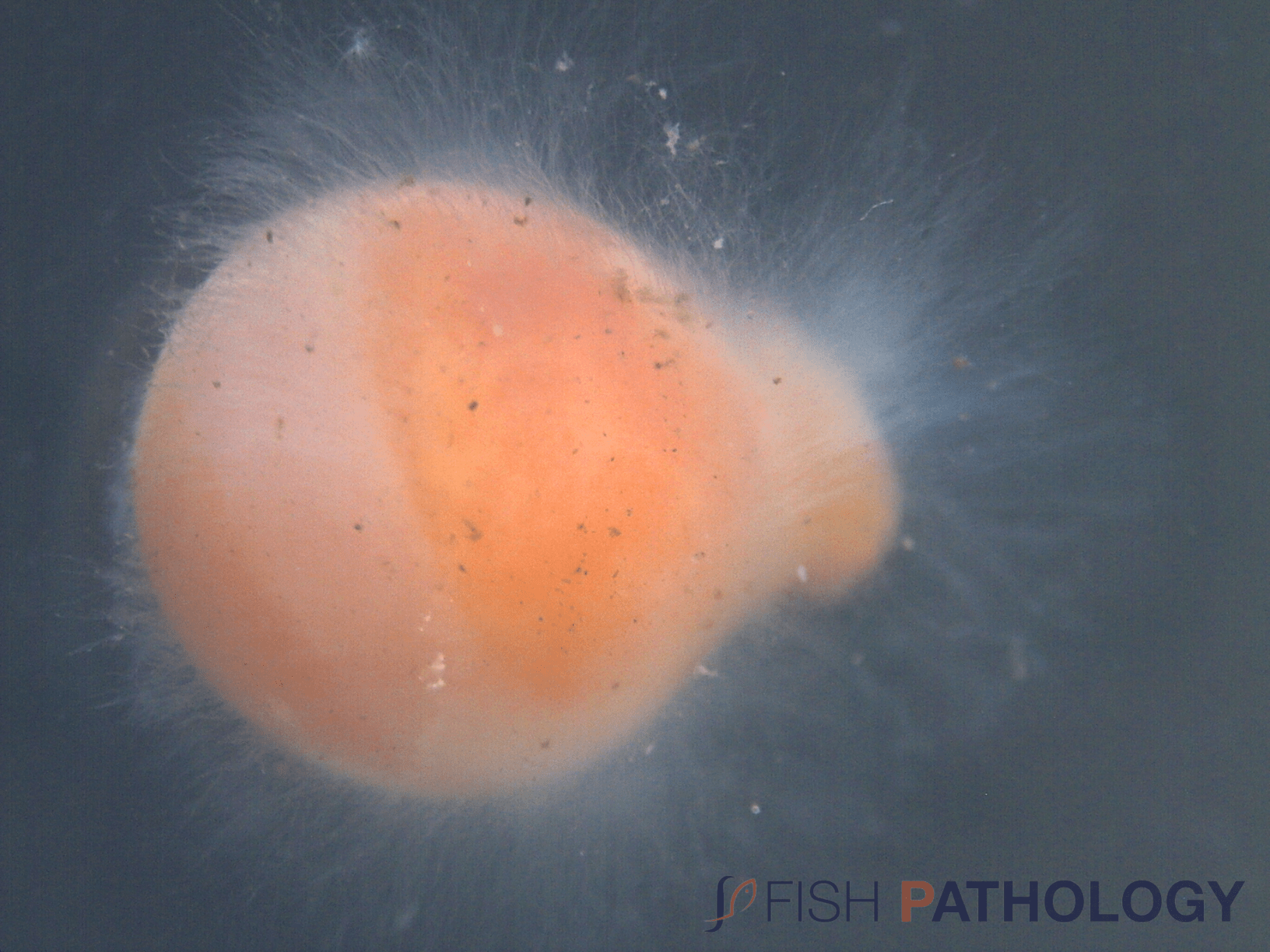
In eggs, mould is detected by the thick layer of mycelium that spreads from dead eggs to healthy eggs, which leads to death by suffocation.
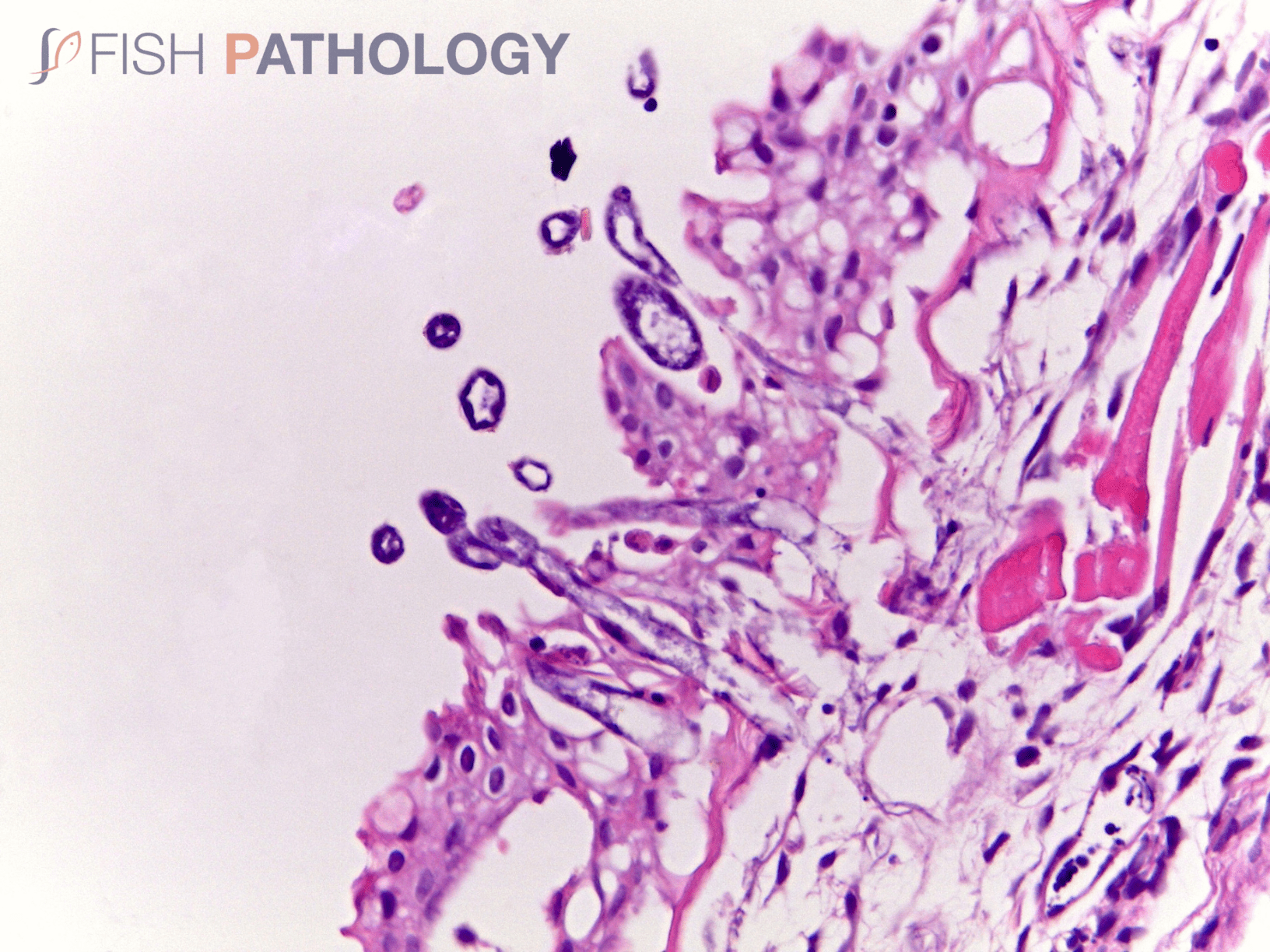
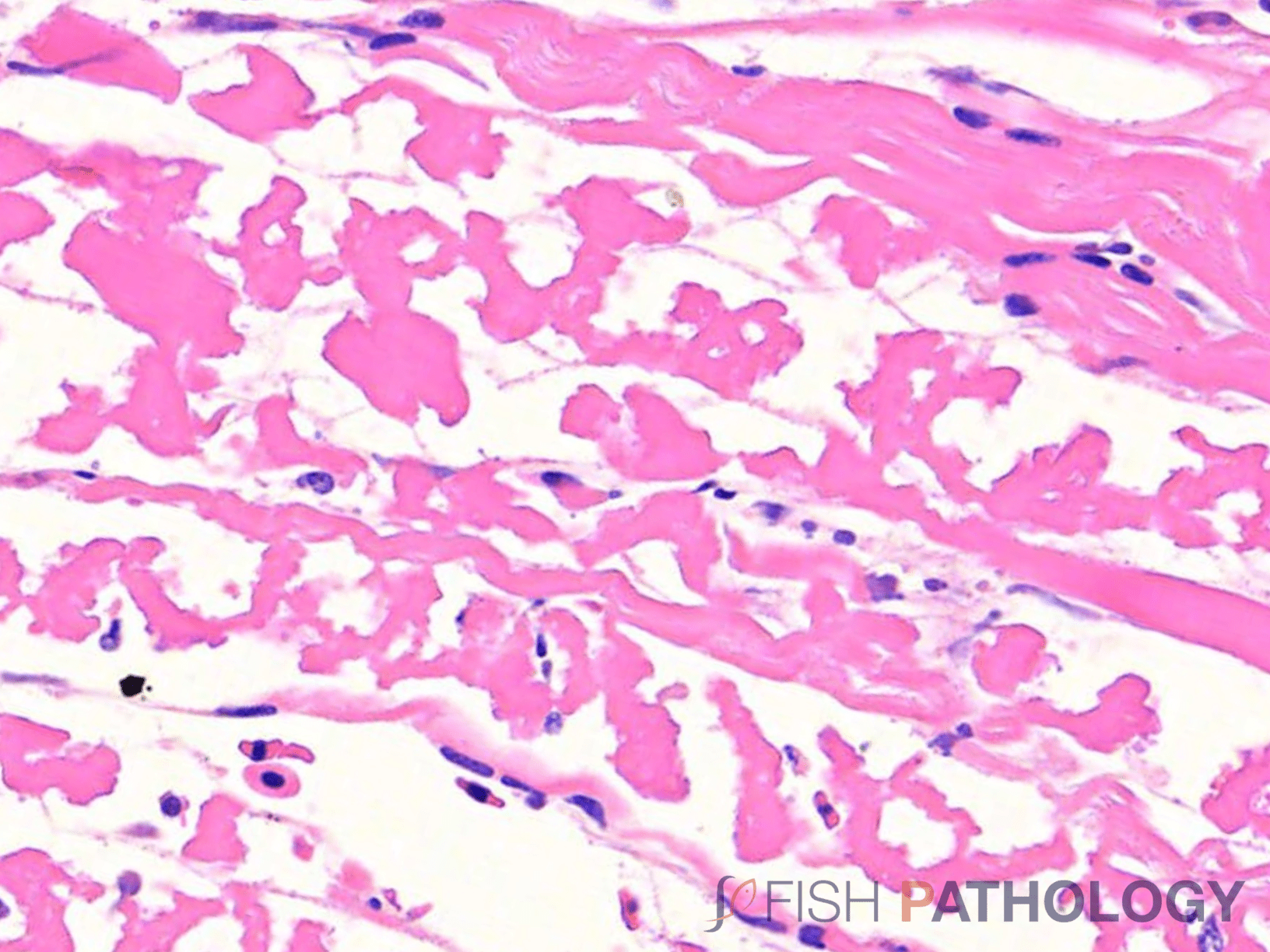
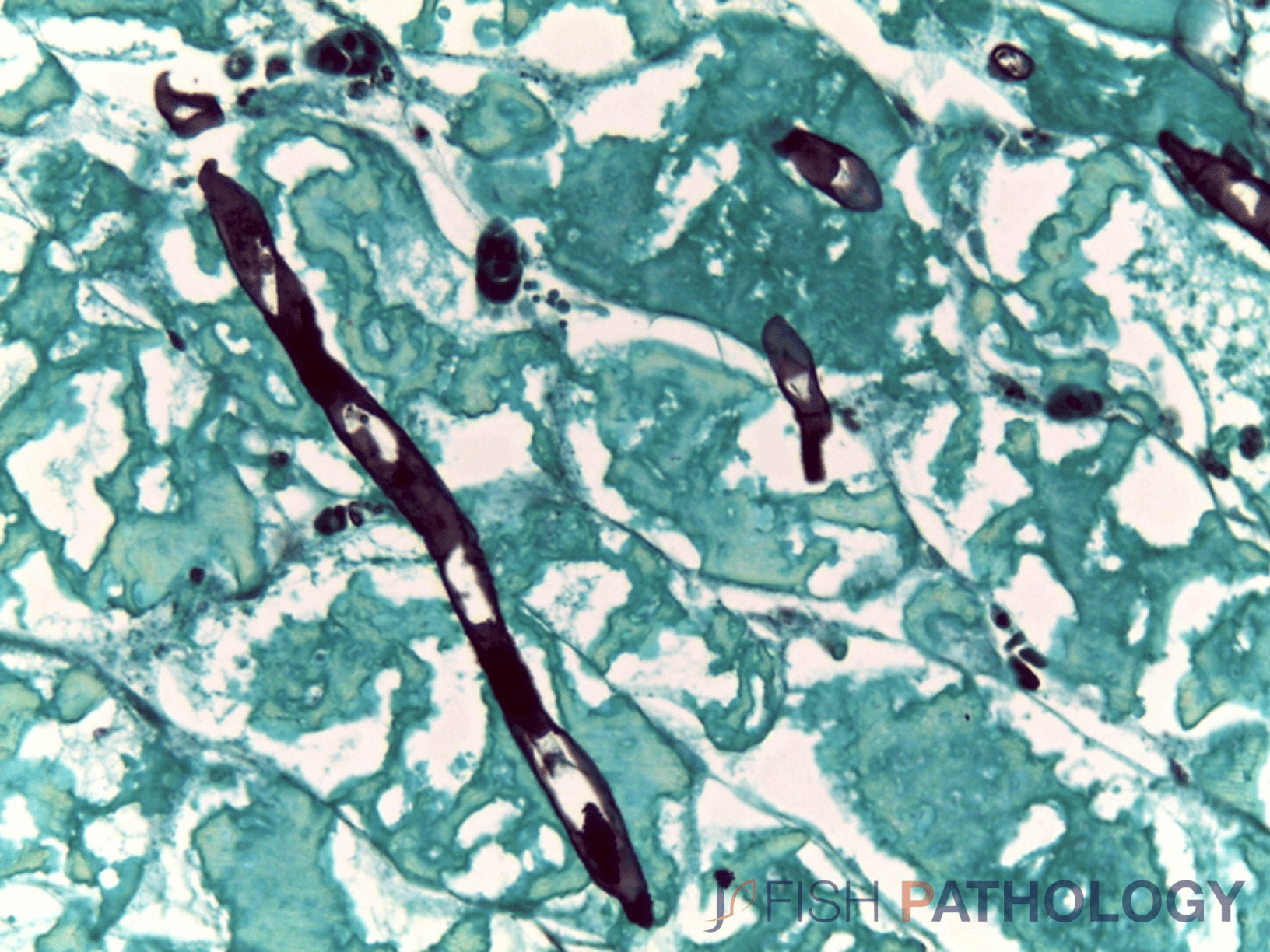
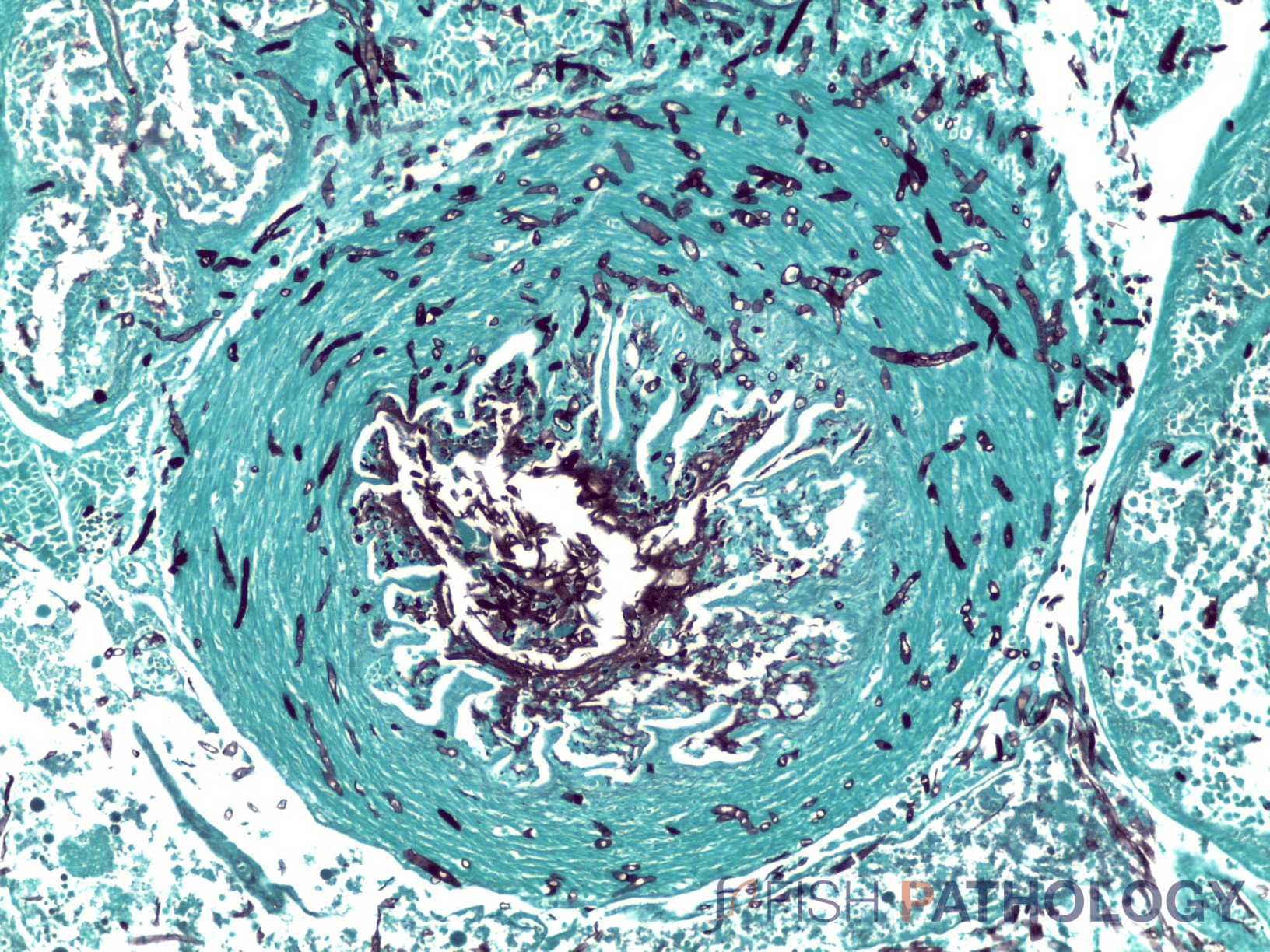
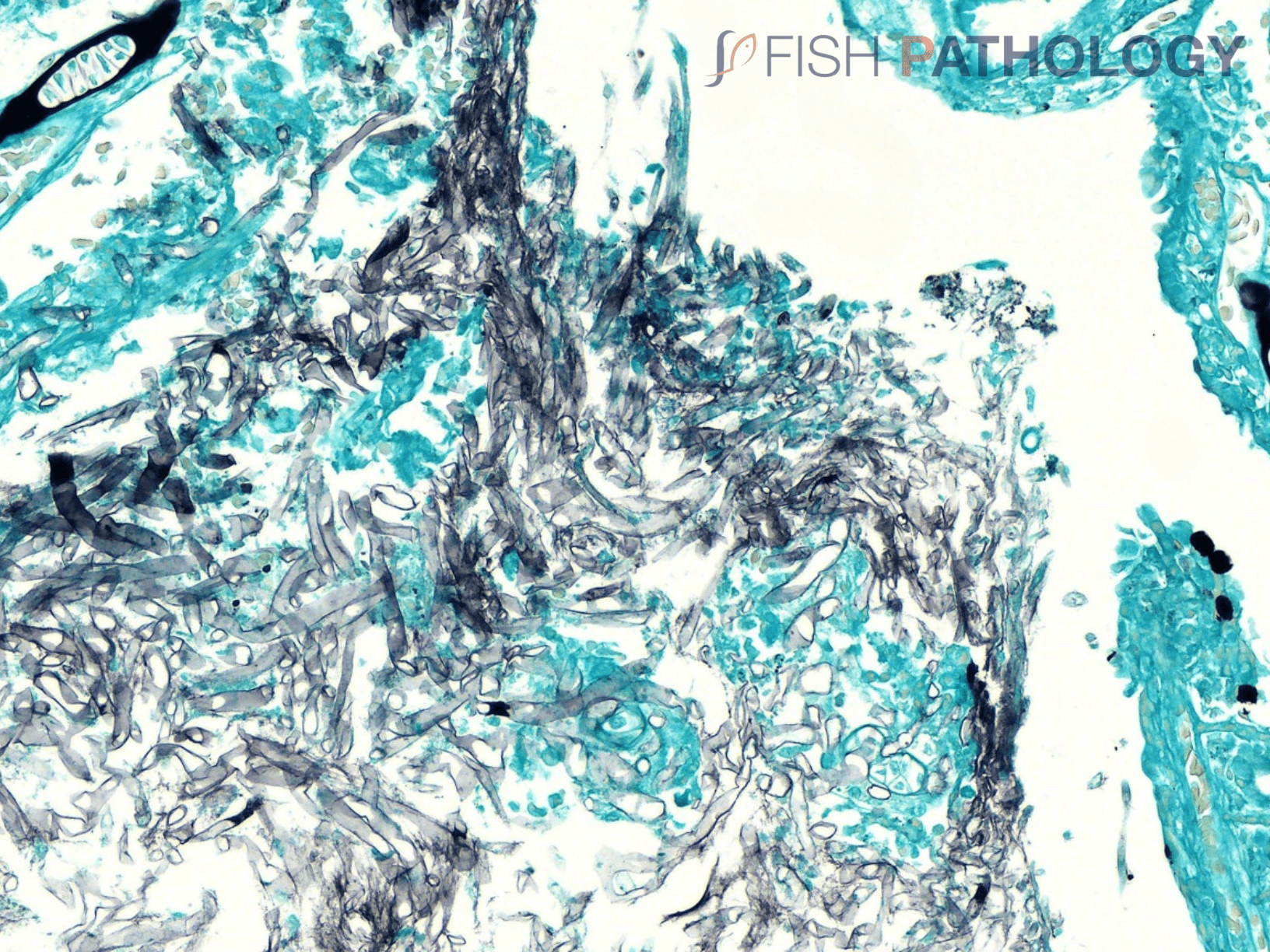
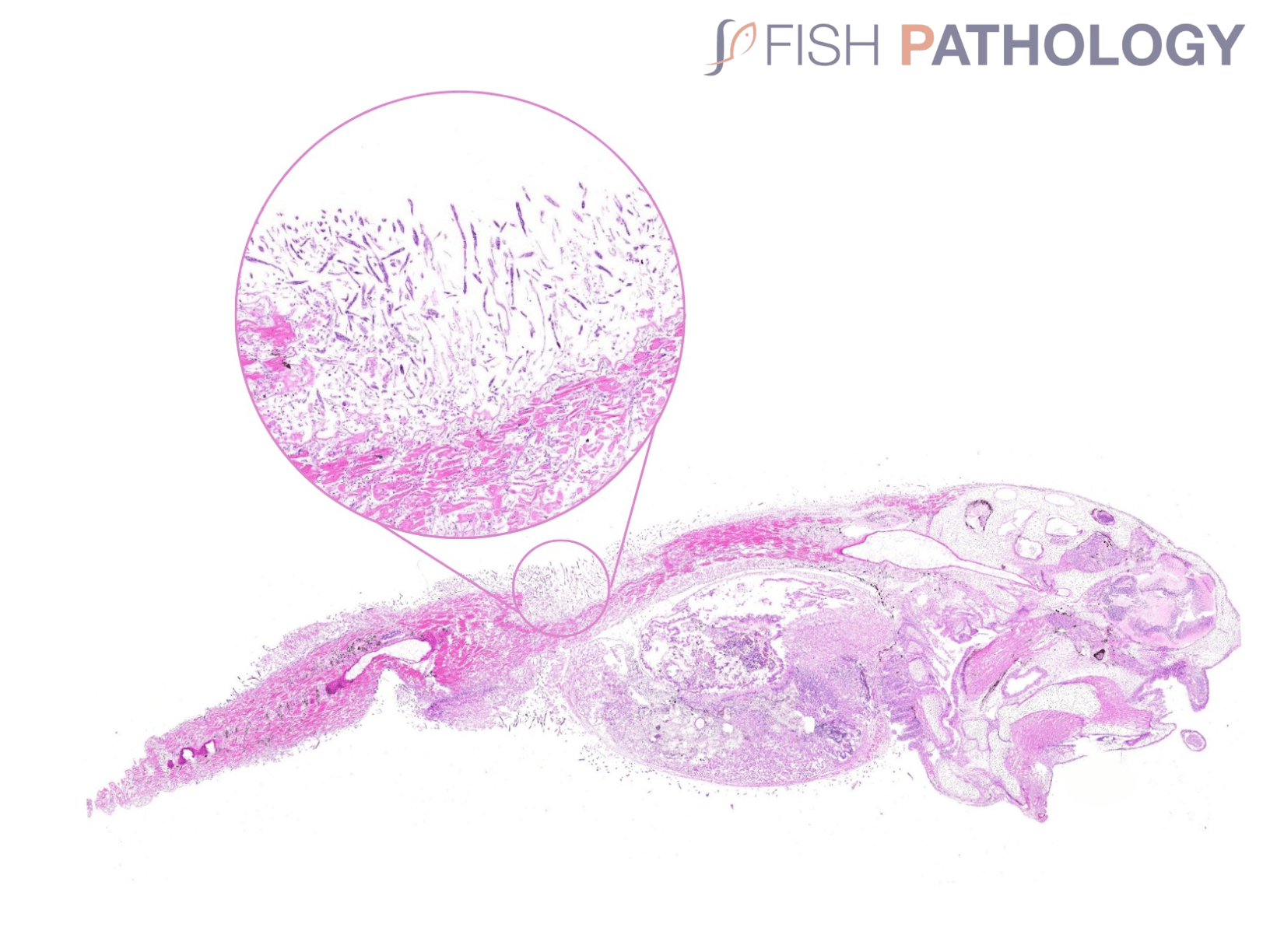
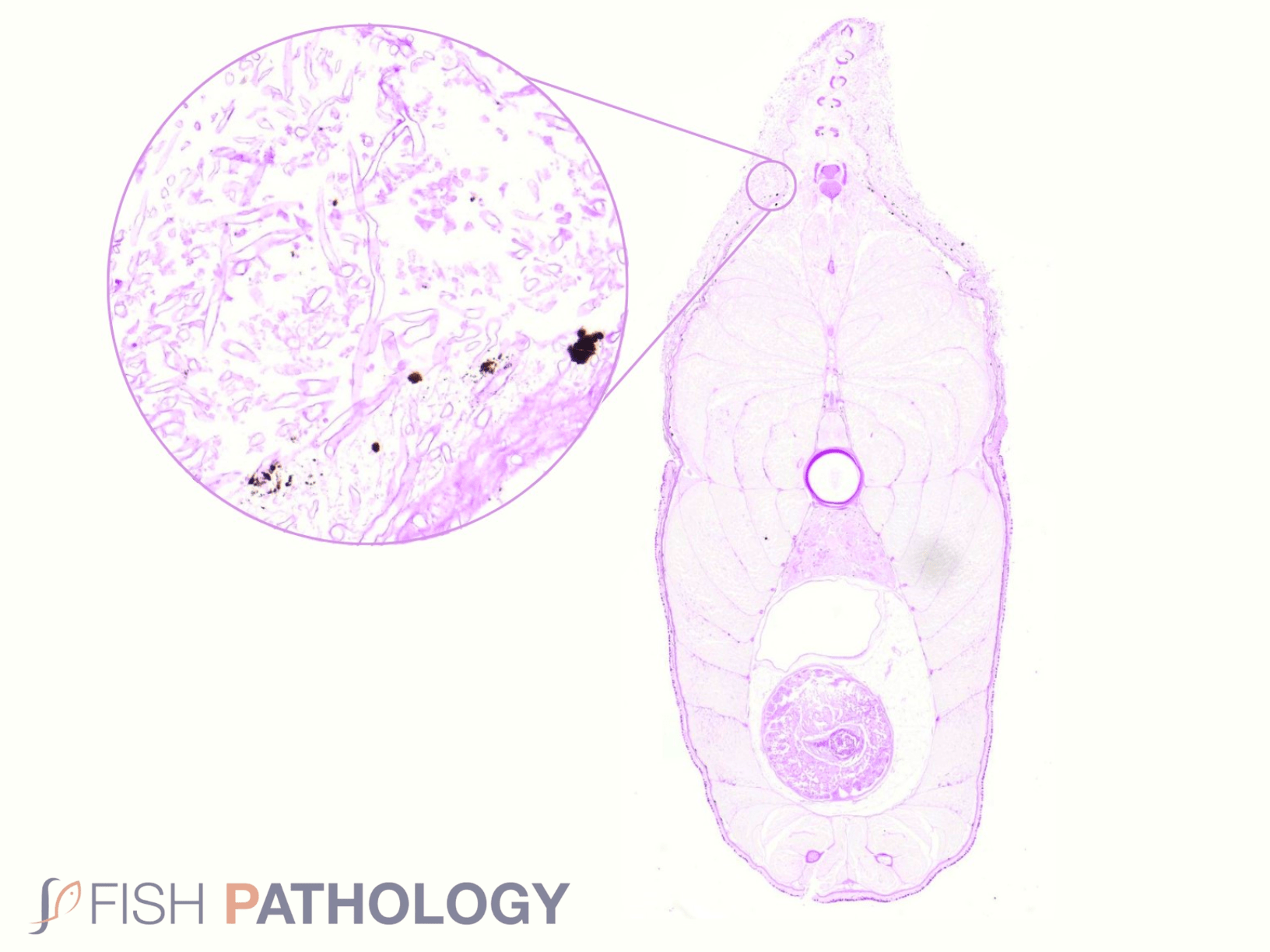
Histologically, the oomycete pathogen usually establishes itself focally, invading the stratum spongiosum of the dermis and then extending over the epidermis, eroding it as it spreads. Superficial invasion of the dermis rapidly leads to osmotic imbalance and peripheral circulatory failure (shock). Also, beneath this superficial mat of mycelium are areas of degenerate tissue ranging from superficial dermal necrosis and oedema to deep myofibrillar necrosis and extensive haemorrhage.
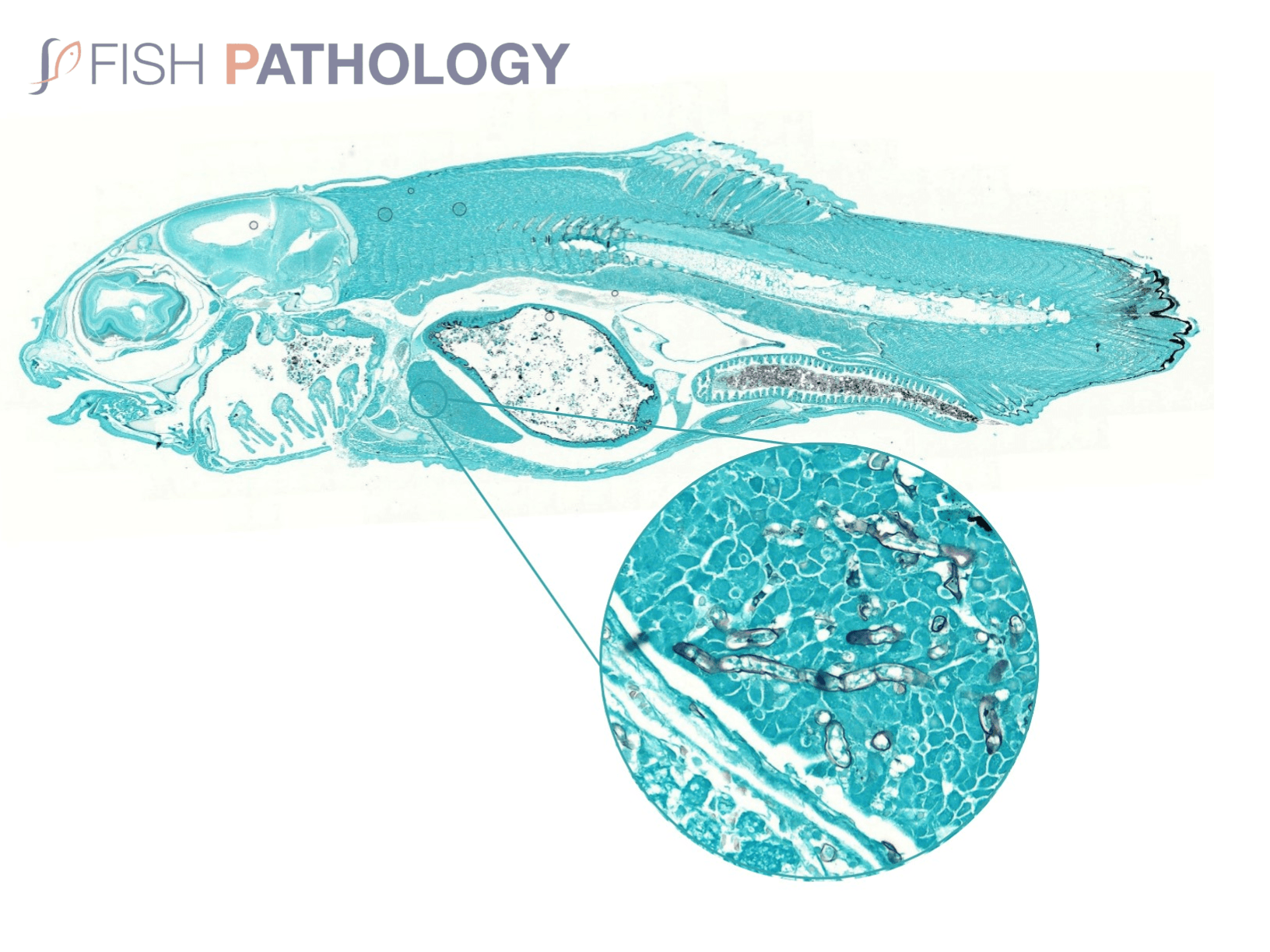
Normally, little inflammation is present, but a more marked response may appear when concomitant bacterial infection occurs. Intestinal infection and peritoneal saprolegniasis are common and non-septate hyphae can be found invading different internal organs such as stomach and kidney. In those cases where invasion is via the gastric lumen with primary mucosal necrosis, likely associations, particularly in larval fish, would be dietary alterations and/or heavy environmental contamination, often due to poor cleaning practices of tanks and feeding equipment.
The oomycete hyphae are PAS-positive and are also easily demonstrated by silver impregnation methods, such as Grocott’s technique.
Images











References
Avendaño-Herrera, R. (2011). Enfermedades Infecciosas del Cultivo de Salmónidos en Chile y el Mundo. 77-104.
Barde, Ravi. (2022). A Review of Saprolegnia Infection on Freshwater Fishes and Control of the Saprolegniosis. International Journal of Health Sciences. 7141-7145.
Beckmann, M. J., Saraiva, M., McLaggan, D., Pottinger, T. G., & van West, P. (2020). Saprolegnia infection after vaccination in Atlantic salmon is associated with differential expression of stress and immune genes in the host. Fish & Shellfish Immunology. Vol. 106, 1095-1105.
Bruno, D. W., Noguera, P. A., & Poppe, T. T. (2013). A Colour Atlas of Salmonid Diseases.
Elameen,A.;Stueland,S.; Kristensen, R.; Fristad, R.F.; Vrålstad, T.; Skaar, I. Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity. J. Fungi 2021,7,713.
Fregeneda-Grandes, J. M., Rodríguez-Cadenas, F., & Aller-Gancedo, J. M. (2007). Fungi isolated from cultured eggs, alevins and broodfish of brown trout in a hatchery affected by saprolegniosis. Journal of Fish Biology, 71(2), 510–518.
Henard, C., Saraiva, M., Ściślak,M., Ruba, T., McLaggan, D., Noguera, P. & van West, P. (2020). Can Ulcerative Dermal Necrosis (UDN) in Atlantic salmon be attributed to ultraviolet radiation and secondary Saprolegnia parasitica infections?Fungal Biology Reviews. Vol. 40, 70-75.
Mastan, S. &. (2018). Observations on saprolegnia infection in freshwater in fishes of Lake Kolleru. International Journal of Fisheries and Aquatic Studies. 6(1), 102-105.
Mustafa, Fadhal. (2020). Influence of phytase enzyme on growth performance and survival rate challenged with saprolegnia spp. In common carp. Iraqi Journal of Agricultural Sciences. 51. 1458-1465.
Noga, E.J. (2000). Fish Disease: Diagnosis and Treatment. Blackwell Publishing Ltd.
Quiniou, S. M.-A., Bigler, S., Clem, L. W., & Bly, J. E. (1998). Effects of water temperature on mucous cell distribution in channel catfish epidermis: a factor in winter saprolegniasis. Fish & Shellfish Immunology, 8(1), 1–11.
Roberts, R.J. (2012). Fish Pathology. Fourth Edition. Blackwell Publishing Ltd, 383-389.
Sandoval-Sierra, J. V., Latif-Eugenin, F., Martín, M. P., Zaror, L., & Diéguez-Uribeondo, J. (2014). Saprolegnia species affecting the salmonid aquaculture in Chile and their associations with fish developmental stage. Aquaculture, 434, 462–469.
Sarowar, M. N., Cusack, R., & Duston, J. (2019). Saprolegnia molecular phylogeny among farmed teleosts in Nova Scotia, Canada. Journal of Fish Diseases.
Scharsack, J. P., & Franke, F. (2022). Temperature effects on teleost immunity in the light of climate change. Journal of fish biology, 1-17.
Stueland, S., Hatai, K., & Skaar, I. (2005). Morphological and physiological characteristics of Saprolegnia spp. strains pathogenic to Atlantic salmon, Salmo salar L. Journal of Fish Diseases, 28(8), 445–453.
Thoen, E., Evensen, Ø., & Skaar, I. (2011). Pathogenicity of Saprolegnia spp. to Atlantic salmon, Salmo salar L., eggs. Journal of Fish Diseases, 34(8), 601–608.
