
Copper is a significant trace element necessary for the normal growth and metabolism of living organisms. However, if there is overexposure, it can be toxic.
Levels of dissolved copper are often increased from anthropogenic origins such as mine washings and direct applications of algicides, molluscicides or antifouling agents.
The most toxic form of copper is the cupric ion (Cu2+). Fish and crustacea are 10 to 100 times more sensitive to the toxicity of copper than mammals. For salmonids, the upper recommended limit is < 0.03 mg/l in hard water (>100 mg/l CaCO3) while in soft water it is <0.0006 mg/l.
In common with many other waterborne pollutants, and in particular with other heavy metals, the primary target of the toxic actions of copper on fish is their gills. High copper concentrations result in rapid mortality, partly a consequence of hypoxaemia, likely due in part to histopathological changes such as cell swelling and thickening of lamellae, congestion, telangiectasia, epithelial detachment, lamellar synechiae and even fusion of lamellae. These changes lead to an overall reduction in gill surface area and impaired laminar water flow, which have the saving effect of reducing exposure to the pollutant. Unfortunately, this very same response impairs oxygen uptake and carbon dioxide excretion, which leads in turn to an increase in PCO2 and acidosis (increased blood lactate). Instead of hypoxaemia, however, mortality likely results from ionoregulatory disturbances that induce elevated blood pressure, leading to cardiovascular collapse.
In addition to impaired gas exchange, copper has also been reported to affect salt balance, acid-base balance, and ammonia excretion, presumably due to the inhibition of Na+ / K+ ATPase, carbonic anhydrase, and transcellular ammonia excretion. There is little evidence to date, however, to support this effect of copper exposure. Additionally, it is suggested that sublethal exposure to copper in marine and freshwater fish induces an increase in plasma cortisol levels, which would cause an increase in protein catabolism, thereby explaining the hyperammonaemia.
Besides disturbing the balance of salt in fish, copper can also have a negative impact on reproductive output at concentrations far below those required to induce mortality or reduce growth. The mechanism by which copper impairs reproduction is unknown but it could be a simple outcome of reduced resource availability arising when energy must be devoted to dealing with copper-induced physiological challenges rather than a direct endocrine effect.
Copper at low concentrations affects hatching. Copper may inactivate chorionase and cause osmotic disturbances that may also affect its activity or the muscular movements necessary to break the eggshell. It has also been associated with low pigmentation of the embryo, spinal cord deformation, cranial malformations, jaw underdevelopment, decreased length, increased time for complete yolk absorption, oedema and opaque yolk sacs.
The impact of copper exposure on the immune system is associated with a decrease in the phagocytic function of macrophages and a decrease in the production of antibodies.
On top of that, it can affect glucose metabolism and cellular structure of fish. Copper exposure causes increased levels of blood glucose, which lead to suppressed appetite and therefore decreased food intake. This process occurs from hormonal changes that increase metabolism of hepatic glycogen into blood glucose (glycogenolysis). All of this influences the high energy demand necessary for metabolic coordination in the liver and thus maintaining the detoxification process in order to overcome chemical stress.
The olfactory system can be also affected. Copper can damage the olfactory epithelium and at the same time reduce the neurophysiological response towards the olfactory stimulant.
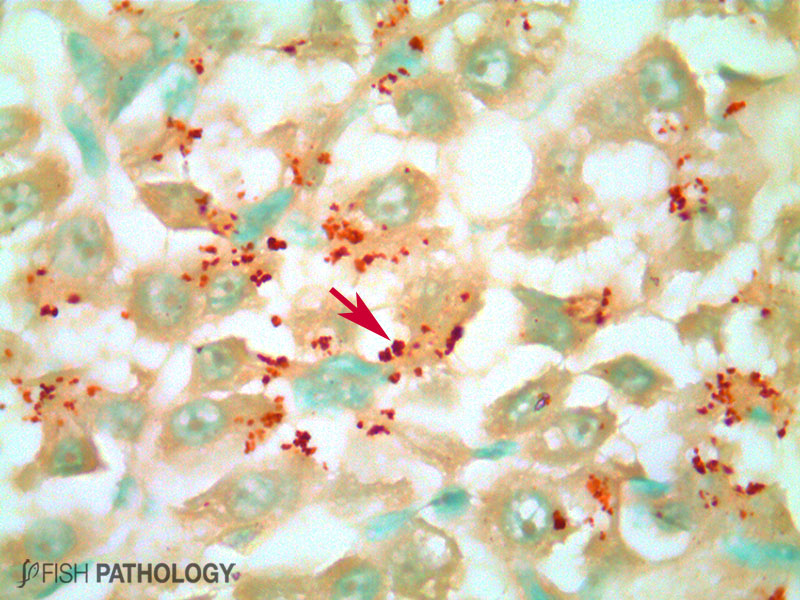
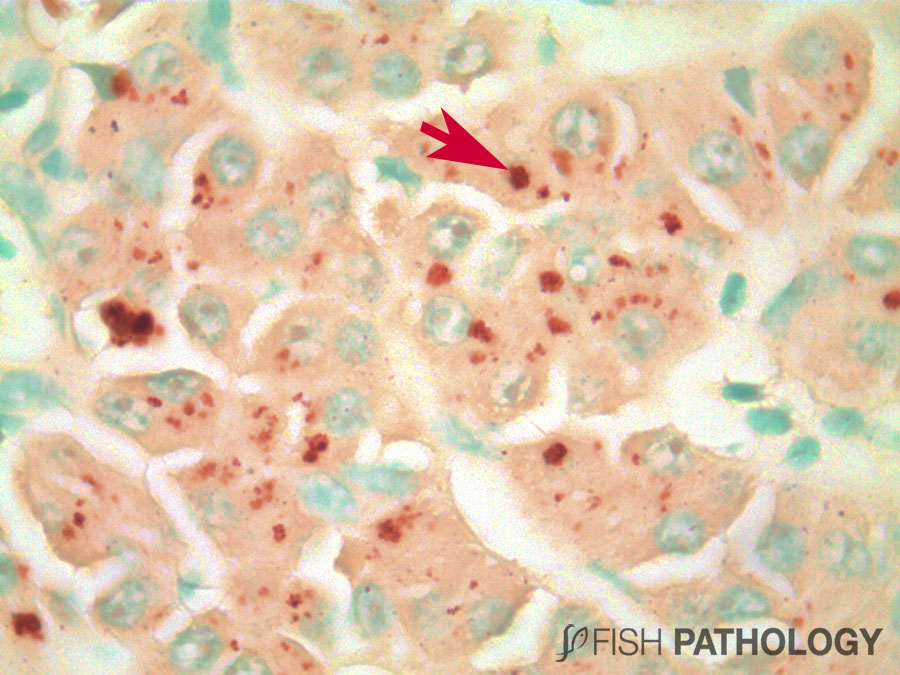
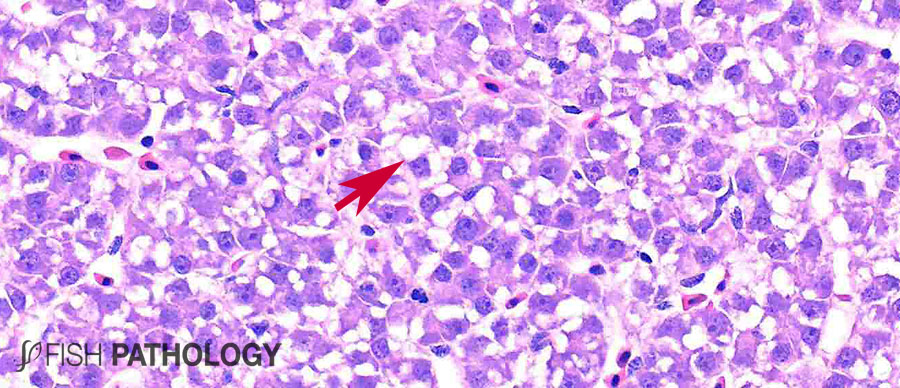
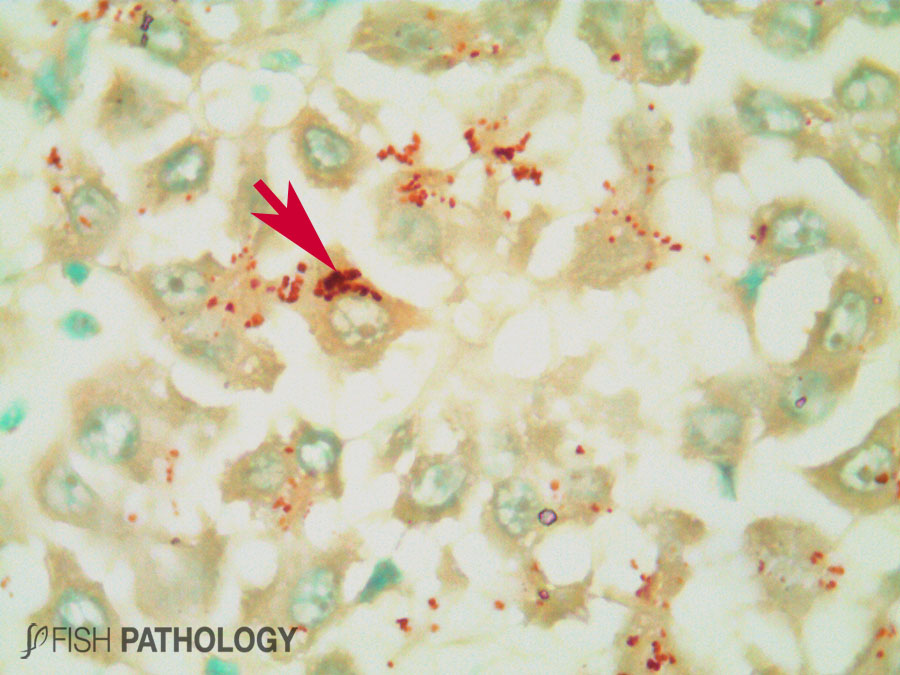
Although both the liver and kidney are typically important for metal accumulation and storage in fish, copper metabolism is primarily controlled by the liver, playing an important role in copper homeostasis. The liver is the most responsive organ to copper accumulation, because it is the place where the detoxification process occurs. Some morphological changes in liver include cytoplasmic vacuolation, dilation of sinusoids, necrosis of hepatic cells, congestion of blood cells and oedema.
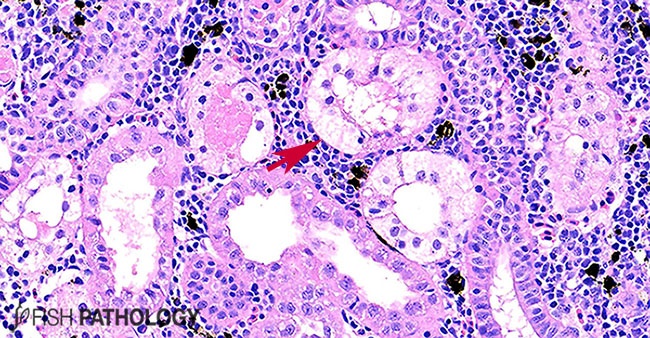
In the kidney, fish can show hydropic degeneration and necrosis of the tubular epithelium, tubular haemorrhage, glomerular shrinkage, collecting duct damage and interstitial congestion.
REFERENCES
- Aghamirkarimi, S., Mashinchian, M. A., Sharifpour, I., Jamili, S., & GHAVAM, M. P. (2017). Sublethal Effects of Copper Nanoparticles on the Histology of Gill, Liver and Kidney of the Caspian Roach, Rutilus Rutilus Caspicus (Case Study).
- Al-Bairuty, G. A., Shaw, B. J., Handy, R. D., & Henry, T. B. (2013). Histopathological effects of waterborne copper nanoparticles and copper sulphate on the organs of rainbow trout (Oncorhynchus mykiss). Aquatic Toxicology, 126, 104-115.
- Di Giulio, R. T., & Hinton, D. E. (Eds.). (2008). The toxicology of fishes. Crc Press.
- Ferguson, Hugh W. (2006). Systemic Pathology of Fish, London, UK, Scotian Press.
- Grosell, M. (2012). Homeostasis and toxicology of essential metals. Fish physiology, 31, 53-133.
- Jezierska, B., Ługowska, K., & Witeska, M. (2009). The effects of heavy metals on embryonic development of fish (a review). Fish physiology and biochemistry, 35(4), 625-640.
- Johnson, A., Carew, E., & Sloman, K. A. (2007). The effects of copper on the morphological and functional development of zebrafish embryos. Aquatic Toxicology, 84(4), 431-438.
- Jones, J., Wellband, K., Zielinski, B., & Heath, D. D. (2019). Transcriptional basis of copper-induced olfactory impairment in the sea lamprey, a primitive invasive fish. G3: Genes, Genomes, Genetics, 9(3), 933-941.
- Mahboob, S., Al-Ghanim, K. A., Al-Balawi, H. F., Al-Misned, F., & Ahmed, Z. (2020). Toxicological effects of heavy metals on histological alterations in various organs in Nile tilapia (Oreochromis niloticus) from freshwater reservoir. Journal of King Saud University-Science, 32(1), 970-973.
- Padrilah, S. N., Shukor, M. Y. A., Yasid, N. A., Ahmad, S. A., Sabullah, M. K., & Shamaan, N. A. (2018). Toxicity Effects of Fish Histopathology on Copper Accumulation. Pertanika Journal of Tropical Agricultural Science, 41(2).
- Sabullah, M. K., Ahmad, S. A., Shukor, M. Y., Gansau, A. J., Syed, M. A., Sulaiman, M. R., & Shamaan, N. A. (2015). Heavy metal biomarker: Fish behavior, cellular alteration, enzymatic reaction and proteomics approaches. International Food Research Journal, 22(2).
- Suchismita, D., & Abhik, G. (2013). Accumulation of copper in different tissues and changes in oxygen consumption rate in Indian flying barb, Esomus danricus (Hamilton-Buchanan) exposed to sub-lethal concentrations of copper. Jordan Journal of Biological Sciences (JJBS), 6(1), 21-24.
- Wood Chris, M., Farrell Anthony, P., & Brauner Colin, J. (2011). Homeostasis and toxicology of essential metals edited. Fish Physiology. London: Academic Press, 31(Part A), 1-497.
