
The negative interactions between jellyfish and fish in aquaculture appear to be an increasing problem. This is partly due to increased numbers of jellyfish, associated with global warming, reduced numbers of their predators, and to the intensification of aquaculture operations in many coastal areas worldwide.
Most reported problems have occurred in marine-farmed salmonids in northwest Europe. Nevertheless, aquaculture operations in other regions such as Asia, North America, and Australia have also been affected.
Jellyfish involved are primarily cnidarians i.e. those species with stinging cells – nematocysts. Several species of jellyfish have been previously linked to fish kill events in marine-farmed fish including hydromedusae, siphonophores, scyphozoans, and ctenophores.
Damage to fish may be direct, through stinging of the skin or gills, or it can be indirect, through de-oxygenation of the surrounding water.
If the species of jellyfish involved is small, it can be washed through the mesh of nets, and even “inhaled” by the fish, thereby damaging the gills. Loose nematocysts from net washing can lead to similar problems. Nematocysts retain their ability to sting long after the jellyfish is dead.
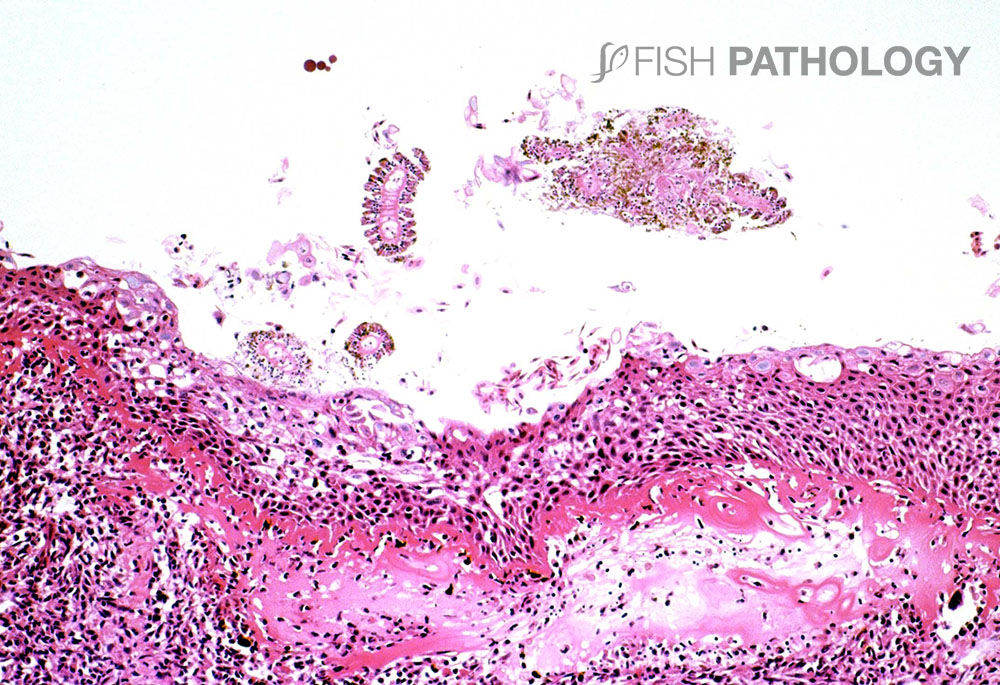
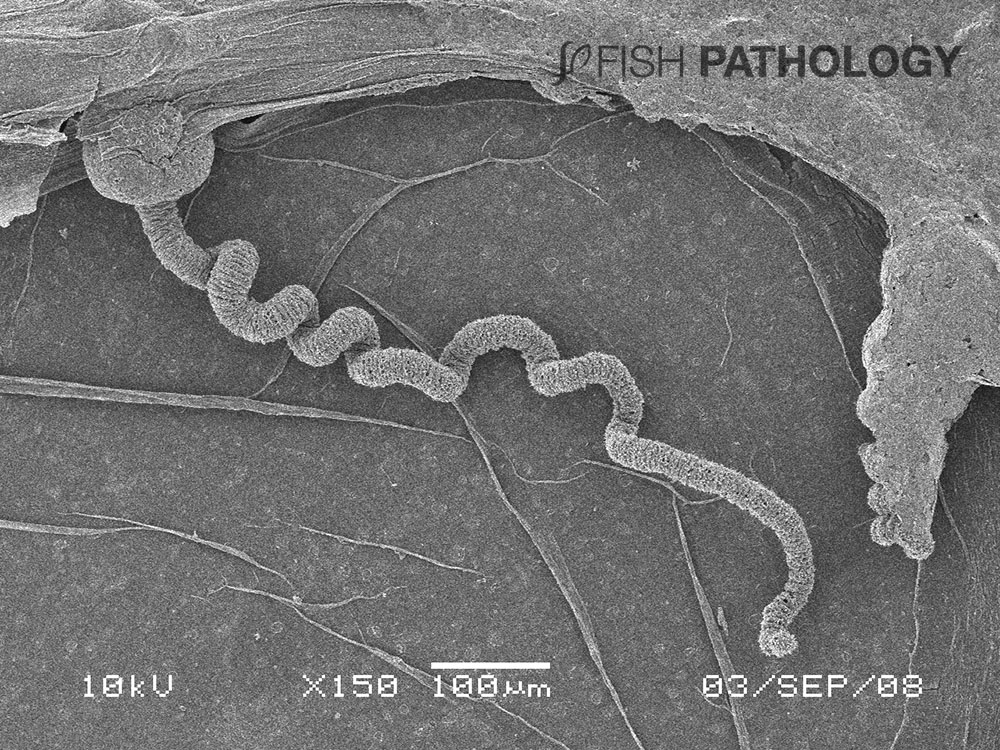
Almost all of the initial gill damage caused by cnidarians is due to their stings. Cnidarian jellyfish are characterized by having millions of microscopic stinging cells, primarily in their tentacles. Inside the cell is a specialized stinging capsule called a nematocyst (mostly 10–20 µm long) that contains a coiled, harpoon-like hollow tube often armed with spines. Nematocyst release can be triggered by mechanical or chemical stimulation. Toxins can be injected from the nematocyst into the prey to immobilize them. When a nematocyst fires, mechanical damage is caused when the tubule penetrates the tissue, rather like a harpoon.
This is followed by toxic damage from the activities of the injected enzymes, neurotoxins, myotoxins, and haemolytic compounds. When the fish does not die directly from the immediate effects of the toxins, it can succumb within a few hours from respiratory failure or later from secondary bacterial infections on the body and the gills caused by opportunistic bacteria such as Tenacibaculum and Vibrio. Some jellyfish have been shown to act as vectors for these bacteria.
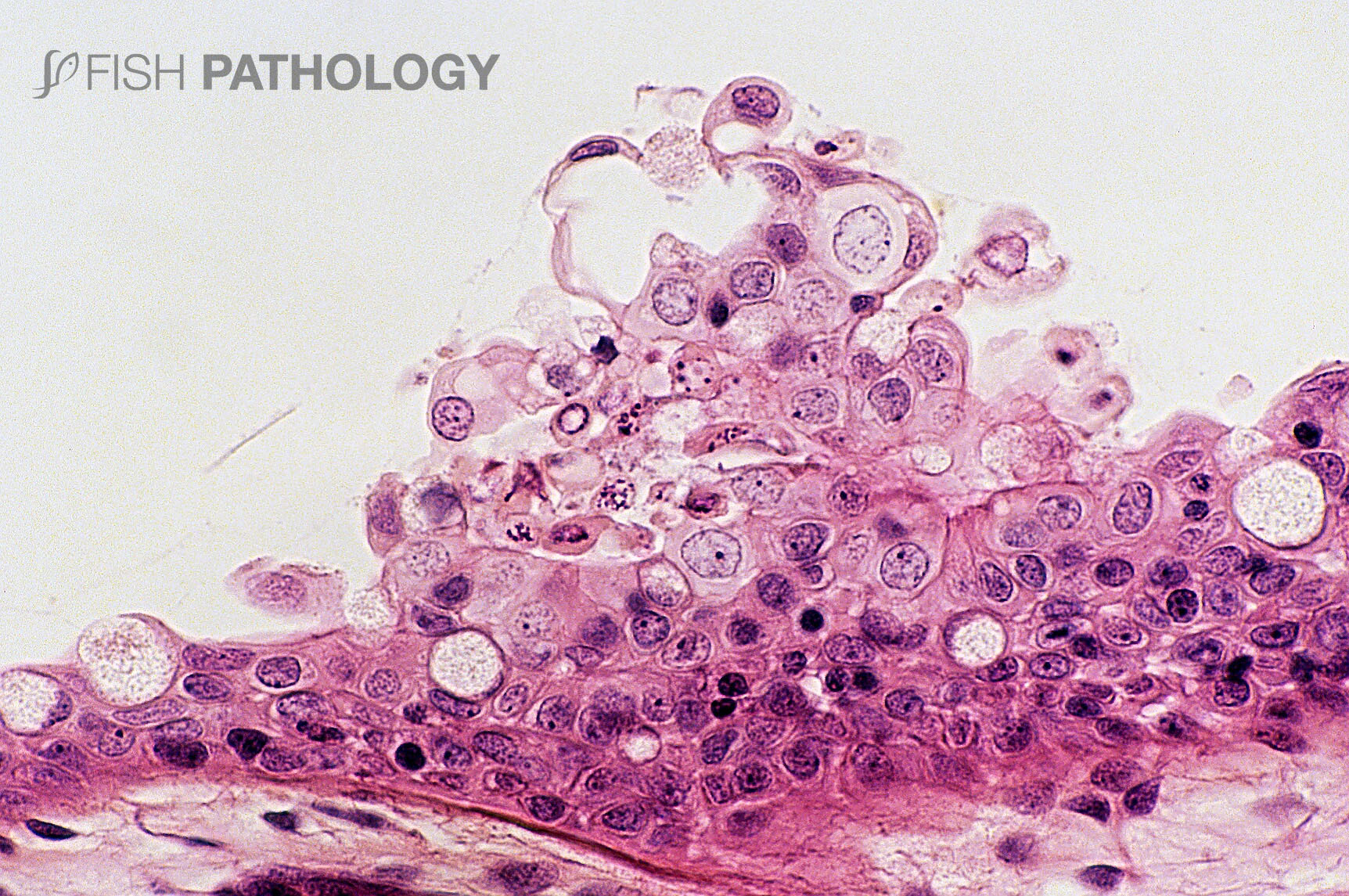
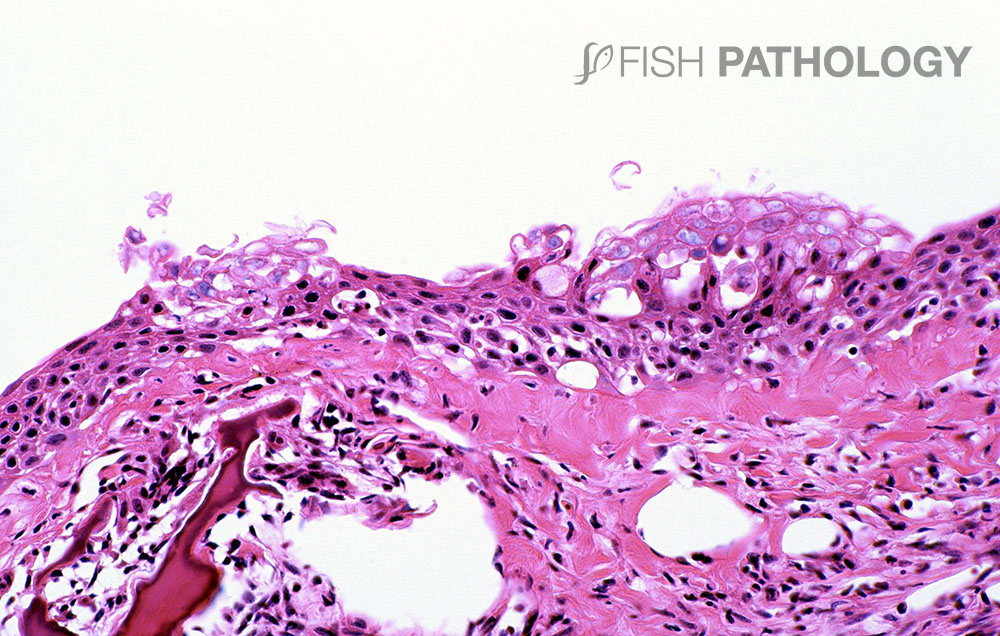
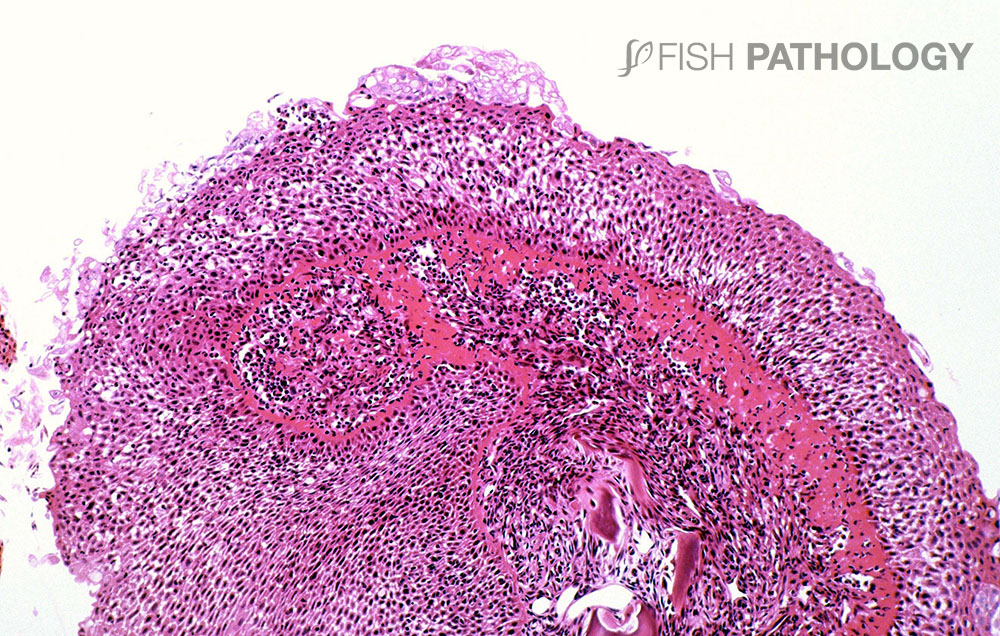
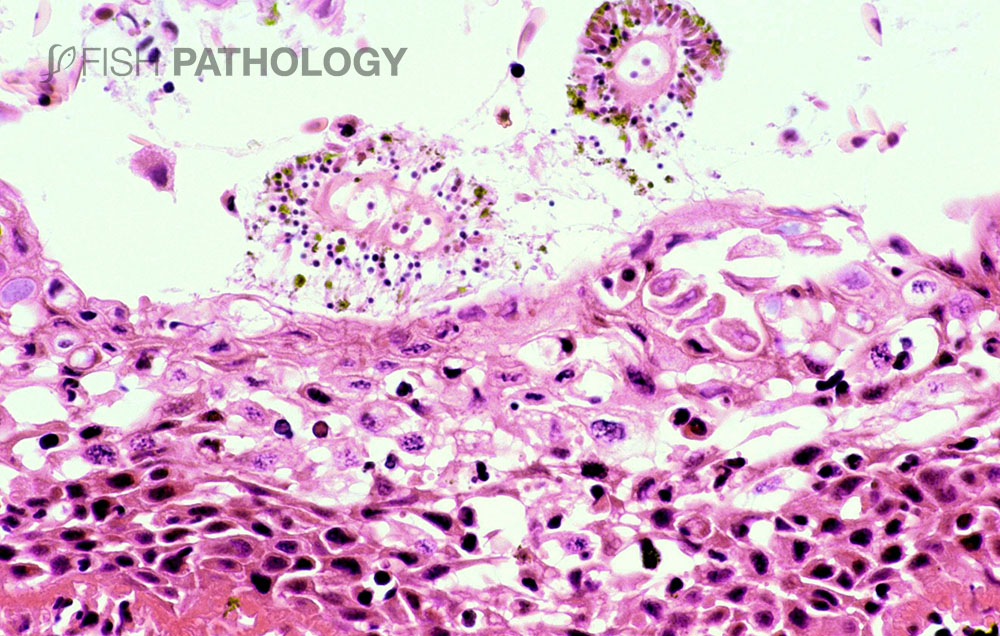
The main organs affected are the gills and skin. Changes to the respiratory epithelium involve lamellar epithelial degeneration and necrosis, exfoliation of epithelial cells, lamellar fusion, congestion, and an infiltration of inflammatory cells, leading to overall lamellar thickening.
On the arch epithelium, superficial lesions are characterized by multifocal acute ballooning degeneration, by spongiosis and hydropic degeneration, and by necrosis.
The inflammatory response is variable but is composed mainly of neutrophils. The presence of marked hyper-eosinophilia of basement membrane and the superficial subepithelial layer, suggest collagen denaturation (necrobiosis). In some cases, the gill rakers are severely necrotic, with sloughing of epithelium, and severe acute inflammation.
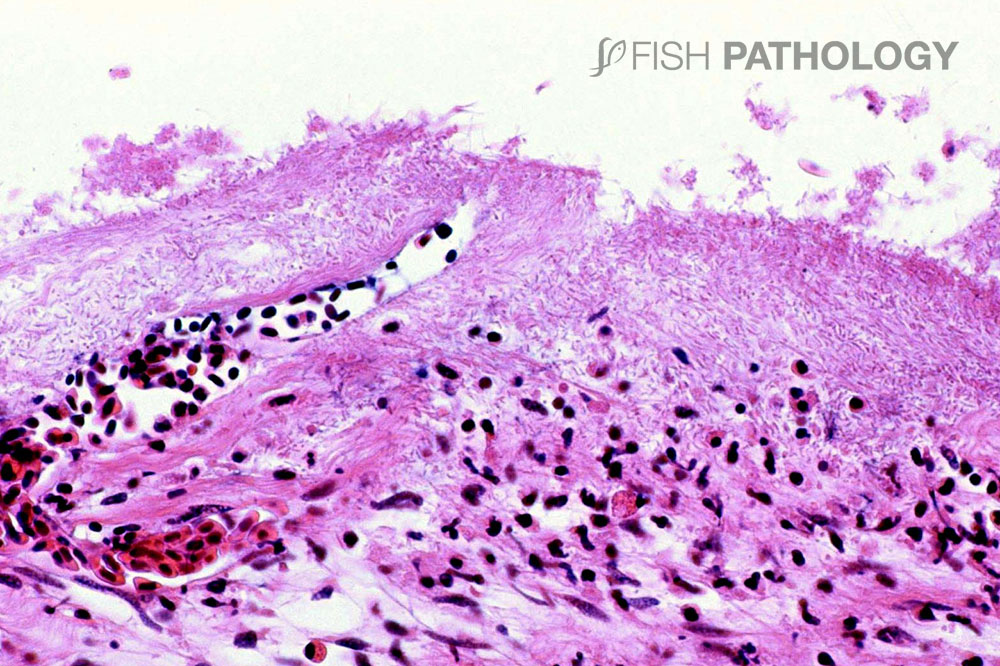
The skin lesions reveal a significant acute dermatitis characterized by a predominantly neutrophilic infiltrate, with the presence of pustule-like aggregates. The dermal stratum spongiosum is usually more affected than the stratum compactum, and haemorrhage, tissue necrosis and oedema can be observed. Generally, the underlying skeletal muscle is not affected.
REFERENCES
- Baxter, E. J., Sturt, M. M., Ruane, N. M., Doyle, T. K., McAllen, R., Harman, L., & Rodger, H. D. (2011). Gill damage to Atlantic salmon (Salmo salar) caused by the common jellyfish (Aurelia aurita) under experimental challenge. PLoS One, 6(4).
- Baxter, E. J., Rodger, H. D., McAllen, R., & Doyle, T. K. (2011). Gill disorders in marine-farmed salmon: investigating the role of hydrozoan jellyfish. Aquaculture Environment Interactions, 1(3), 245-257.
- Ferguson, H. W., Christian, M. D., Hay, S., Nicolson, J., Sutherland, D., & Crumlish, M. (2010). Jellyfish as vectors of bacterial disease for farmed salmon (Salmo salar). Journal of veterinary diagnostic investigation, 22(3), 376-382.
- Marcos-López, M., Mitchell, S. O., & Rodger, H. D. (2016). Pathology and mortality associated with the mauve stinger jellyfish Pelagia noctiluca in farmed Atlantic salmon Salmo salar L. J. Fish Dis, 39, 111-115.
- Purcell, J. E., Baxter, E. J., & Fuentes, V. L. (2013). Jellyfish as products and problems of aquaculture. In Advances in aquaculture hatchery technology (pp. 404-430). Woodhead Publishing.
